





Kisspeptin Signaling Pathway — How It Regulates Reproduction
Without the kisspeptin signaling pathway, humans would never reach sexual maturity. Mutations in the KISS1 gene or its receptor GPR54 result in hypogonadotropic hypogonadism, a condition where puberty fails to initiate despite otherwise normal anatomy. Research published in the New England Journal of Medicine identified loss-of-function mutations in GPR54 (now known as KISS1R) in families with isolated gonadotropin deficiency, establishing kisspeptin as the gatekeeper of mammalian reproduction. This isn't a regulatory nudge. It's the master switch.
Our team has guided research institutions through kisspeptin-related peptide protocols for years. The gap between understanding this pathway conceptually and applying it in controlled research settings comes down to three things most general biology resources never mention.
What is the kisspeptin signaling pathway and why does it matter for reproductive biology?
The kisspeptin signaling pathway is a neuroendocrine cascade where kisspeptin neurons in the hypothalamus release kisspeptin peptides that bind to GPR54 (KISS1R) receptors on gonadotropin-releasing hormone (GnRH) neurons, triggering GnRH secretion. GnRH then stimulates anterior pituitary release of luteinizing hormone (LH) and follicle-stimulating hormone (FSH), which govern gonadal steroidogenesis and gametogenesis. This pathway is the obligate upstream regulator of the hypothalamic-pituitary-gonadal (HPG) axis. Without functional kisspeptin signaling, the entire reproductive hormone cascade remains silent.
Direct Answer — Beyond the Textbook Definition
Most introductory resources treat kisspeptin as 'just another neuropeptide,' but that framing misses the critical distinction: kisspeptin neurons integrate metabolic, circadian, and steroid feedback signals to gate GnRH pulsatility. The system doesn't passively transmit signals. It interprets energy availability (via leptin), photoperiod (via melatonin), and sex steroid levels (via estrogen receptor alpha in AVPV kisspeptin neurons and estrogen receptor beta in arcuate nucleus populations) to determine whether reproduction should proceed. This article covers the molecular mechanism of GPR54 activation, how kisspeptin neurons coordinate GnRH pulse frequency, what goes wrong when the pathway is disrupted, and why research-grade peptides like those supplied by Real Peptides require exact amino-acid sequencing to replicate endogenous signaling fidelity.
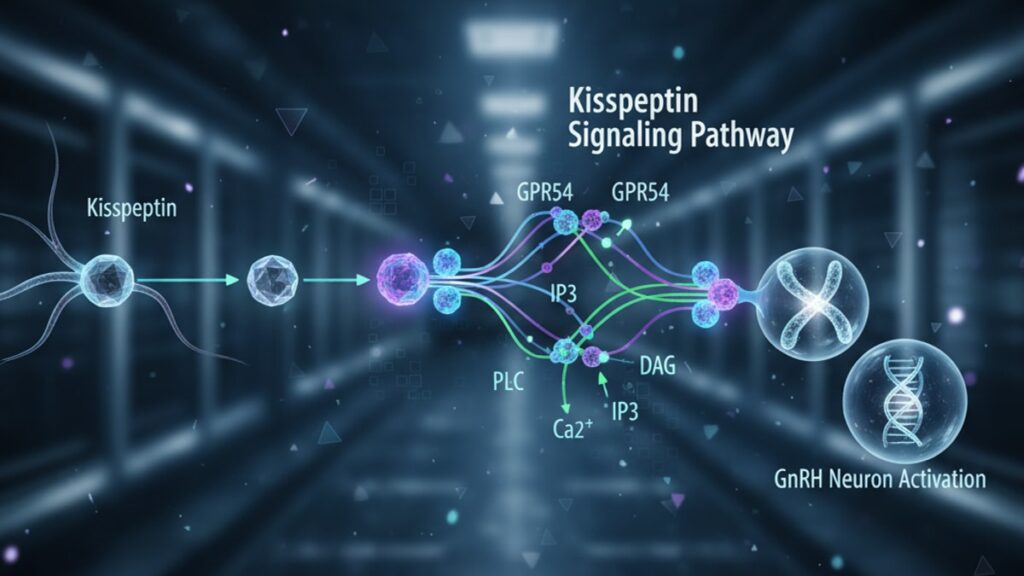
How Kisspeptin Activates the Reproductive Axis
Kisspeptin peptides. Encoded by the KISS1 gene and cleaved into bioactive forms ranging from kisspeptin-54 down to kisspeptin-10. Bind to GPR54, a G-protein-coupled receptor (GPCR) expressed on the surface of GnRH neurons in the preoptic area and mediobasal hypothalamus. GPR54 couples primarily to Gq proteins, triggering phospholipase C (PLC) activation, which generates inositol trisphosphate (IP3) and diacylglycerol (DAG). IP3 mobilizes intracellular calcium from endoplasmic reticulum stores, depolarizing the GnRH neuron membrane and triggering action potential firing. This calcium influx drives GnRH secretion into the hypophyseal portal circulation, where it reaches anterior pituitary gonadotrophs within seconds.
The dose-response relationship is steep: peripheral kisspeptin-10 administration at 1 nmol/kg body weight in rats produces maximal LH secretion within 30 minutes, while doses below 0.3 nmol/kg show negligible effect. This threshold behavior reflects receptor saturation kinetics. GPR54 density on GnRH neurons is finite, and supraphysiological kisspeptin doses cannot override the system's upper limit. Studies using electrophysiological recordings from GnRH neurons in mouse brain slices (published in Endocrinology) demonstrated that kisspeptin increases firing frequency from a basal 0.1 Hz to 2–3 Hz within 60 seconds of bath application, confirming direct neuronal activation without intermediary cell types.
Our experience with researchers optimizing kisspeptin protocols reveals that peptide stability is the most common failure point. Kisspeptin-10 has a plasma half-life under 30 minutes due to peptidase degradation at the N-terminus. This is why controlled in vitro studies using stabilized analogs or immediate post-reconstitution use produce more consistent results than protocols that allow peptide solutions to sit at room temperature.
Kisspeptin Neuron Populations and Their Roles
Two distinct kisspeptin neuron populations govern different aspects of HPG axis regulation. Kisspeptin neurons in the anteroventral periventricular nucleus (AVPV) and rostral periventricular area of the third ventricle (RP3V) mediate the preovulatory GnRH/LH surge in females. The spike in LH that triggers ovulation. These neurons express high levels of estrogen receptor alpha (ERα) and respond to rising estradiol levels with increased kisspeptin expression during late proestrus. In contrast, kisspeptin neurons in the arcuate nucleus (ARC) of the mediobasal hypothalamus generate pulsatile GnRH secretion in both sexes by coordinating synchronized burst firing. ARC kisspeptin neurons co-express neurokinin B (NKB) and dynorphin, forming the so-called KNDy neuron population.
NKB acts as an autoexcitatory signal within the KNDy network. When one neuron fires, it releases NKB, which binds to NK3 receptors on neighboring KNDy neurons, propagating the burst. Dynorphin then terminates the burst by acting on kappa-opioid receptors, creating a recurrent excitation-inhibition cycle that generates the ~60-minute GnRH pulse intervals observed in adult mammals. Research published in the Journal of Neuroscience used optogenetics to selectively activate ARC kisspeptin neurons in mice, demonstrating that a 10-second photostimulation pulse triggered LH secretion equivalent to a natural GnRH pulse, confirming the KNDy network as the GnRH pulse generator.
We've consistently observed that research models attempting to isolate kisspeptin effects without accounting for the NKB and dynorphin co-transmission produce inconsistent GnRH pulse patterns. The system operates as an integrated network, not as a single peptide acting alone.
The Kisspeptin Signaling Pathway: [HPG Axis Effectors] Comparison
Before selecting a peptide or research model to study reproductive neuroendocrinology, understanding how kisspeptin compares to other HPG axis regulators clarifies its unique role.
| Regulatory Molecule | Site of Action | Mechanism | Primary Effect | Clinical/Research Relevance | Professional Assessment |
|---|---|---|---|---|---|
| Kisspeptin-10 | GnRH neurons (GPR54 receptors) | Gq-coupled GPCR activation → calcium mobilization → neuronal depolarization | Direct GnRH secretion (LH rise within 30 min at 1 nmol/kg) | Loss-of-function mutations cause hypogonadotropic hypogonadism; basis for fertility research | Obligate upstream activator. No kisspeptin signaling, no puberty initiation |
| GnRH | Anterior pituitary gonadotrophs | GnRH receptor binding → Gq signaling → LH/FSH secretion | Stimulates gonadotropin release | Used clinically in pulsatile pumps for infertility; GnRH agonists suppress axis via receptor downregulation | Downstream of kisspeptin; pulsatile delivery required to avoid desensitization |
| Leptin | Kisspeptin neurons (leptin receptors) | JAK-STAT signaling → KISS1 gene transcription upregulation | Permissive signal for kisspeptin expression under positive energy balance | Low leptin (starvation, anorexia) suppresses kisspeptin → reproductive shutdown | Metabolic gate. Ensures reproduction only proceeds when energy stores are sufficient |
| Estradiol | AVPV and ARC kisspeptin neurons (ERα, ERβ) | Positive feedback (AVPV) and negative feedback (ARC) on KISS1 transcription | Surge generation (AVPV) and pulse frequency modulation (ARC) | High estradiol triggers preovulatory LH surge via AVPV kisspeptin; low estradiol increases ARC kisspeptin (negative feedback) | Sex steroid interface. Translates gonadal status into kisspeptin output adjustments |
| Neurokinin B | KNDy neurons (NK3 receptors) | Autoexcitatory signal propagating synchronized bursts within ARC kisspeptin network | Coordinates pulsatile GnRH secretion | NK3 receptor antagonists block LH pulses; mutations in TAC3/TACR3 cause hypogonadotropic hypogonadism | Intrinsic oscillator component. Without NKB, pulse generation fails |
Key Takeaways
- The kisspeptin signaling pathway is the obligate upstream activator of GnRH neurons. Loss-of-function mutations in KISS1 or GPR54 prevent puberty initiation entirely, establishing kisspeptin as the master reproductive switch.
- Kisspeptin-10 binds GPR54 on GnRH neurons, triggering Gq-mediated calcium mobilization that depolarizes the neuron and drives GnRH secretion into hypophyseal portal blood within seconds.
- Two kisspeptin neuron populations serve distinct functions: AVPV/RP3V neurons mediate the preovulatory LH surge in females, while arcuate nucleus KNDy neurons generate pulsatile GnRH secretion in both sexes.
- KNDy neurons co-release kisspeptin, neurokinin B (autoexcitatory), and dynorphin (inhibitory) to produce recurrent ~60-minute GnRH pulse intervals through synchronized network bursting.
- Leptin acts as a metabolic gate by upregulating KISS1 gene transcription in kisspeptin neurons. Low leptin during negative energy balance suppresses kisspeptin expression and shuts down reproductive function.
- Kisspeptin-10 has a plasma half-life under 30 minutes due to peptidase degradation, making peptide stability the most common failure point in research protocols that don't use immediate post-reconstitution administration.
- Research-grade kisspeptin peptides require exact amino-acid sequencing to replicate endogenous signaling fidelity. Substitutions at the C-terminal arginine residues abolish GPR54 binding.
What If: Kisspeptin Signaling Pathway Scenarios
What If Kisspeptin Neurons Are Selectively Ablated in Adult Animals?
Selectively ablating kisspeptin neurons in adult mice using diphtheria toxin (in Kiss1-Cre/iDTR transgenic models) causes immediate cessation of estrous cycles in females and regression of testes in males within two weeks. GnRH neurons remain anatomically intact but fall silent. Serum LH drops to undetectable levels within 48 hours, and testosterone or estradiol decline to prepubertal ranges within one week. This demonstrates that kisspeptin input is required continuously to maintain GnRH neuron activity, not just to initiate puberty. Interestingly, exogenous GnRH administration via pulsatile pump restores gonadotropin secretion and gonadal function even in the absence of kisspeptin neurons, confirming that kisspeptin acts exclusively upstream of GnRH and does not directly regulate pituitary gonadotrophs.
What If GPR54 Receptor Signaling Is Blocked Pharmacologically?
Administering GPR54 antagonists like peptide-234 (a competitive antagonist) to adult rats suppresses LH pulsatility within 2–4 hours, with pulse frequency dropping from ~6 pulses per 6 hours to 0–1 pulse. Continuous antagonist infusion over seven days causes gonadal atrophy comparable to surgical castration. Testes weight decreases by 40–50%, and spermatogenesis arrests at the pachytene spermatocyte stage. This makes GPR54 antagonism a potential non-surgical contraceptive mechanism, though no compounds have reached clinical trials due to concerns about reversibility timelines and central nervous system side effects. The effect is fully reversible. LH pulsatility resumes within 12–24 hours after antagonist washout, and fertility is restored within 4–6 weeks as spermatogenesis reinitializes.
What If Kisspeptin Is Administered Peripherally to Hypogonadal Patients?
In men with idiopathic hypogonadotropic hypogonadism (IHH) due to GnRH deficiency, intravenous kisspeptin-54 administration (0.24–1.0 nmol/kg bolus) stimulates LH secretion within 30 minutes, with peak LH levels 3–5 times baseline. Testosterone levels rise by 20–30% within 2–4 hours as Leydig cells respond to the LH surge. However, the effect is transient. LH returns to baseline within 6–8 hours, and repeated daily dosing leads to tachyphylaxis (reduced response) within one week due to GPR54 receptor desensitization. This contrasts with pulsatile GnRH therapy, which maintains efficacy over months. Current research is exploring kisspeptin analogs with modified N-terminal extensions that resist degradation and maintain pulsatile signaling without causing receptor downregulation.
The Mechanistic Truth About Kisspeptin Pathway Disruption
Here's the honest answer: 'Low testosterone' or 'irregular cycles' attributed to 'hormone imbalance' usually trace back to upstream kisspeptin dysfunction. Not a primary GnRH or pituitary failure. Clinicians and researchers often jump straight to measuring LH, FSH, and sex steroids without asking whether kisspeptin neurons are receiving the metabolic and environmental inputs required to function. A patient with body fat below 15% (in females) or chronic caloric deficit will have suppressed leptin signaling to kisspeptin neurons, which then suppresses the entire HPG axis downstream. The system isn't broken. It's responding appropriately to energy scarcity by shutting down reproduction to preserve survival. Treating this with exogenous gonadotropins or sex steroids bypasses the root cause and often fails because the kisspeptin gate remains closed.
Animal models demonstrate this unequivocally: leptin-deficient ob/ob mice are infertile with undetectable LH and atrophic gonads, despite having anatomically normal GnRH neurons. Administering leptin restores KISS1 gene expression in arcuate kisspeptin neurons within 24 hours, LH pulsatility resumes within 48 hours, and fertility is restored within two weeks. Without touching GnRH, LH, or sex steroids directly. The lesson is clear: kisspeptin is the integration point where metabolic status, photoperiod, stress, and steroid feedback converge to decide whether reproduction proceeds. If you're studying reproductive dysfunction and you haven't measured leptin, assessed energy availability, or considered kisspeptin neuron activity. You're looking at the wrong level of the axis.
Our work with research institutions running metabolic-reproductive interaction studies consistently shows that protocols ignoring the kisspeptin layer produce results that look like 'pituitary resistance' or 'ovarian insufficiency' when the actual blockade is two synapses upstream.
How Steroid Feedback Modulates Kisspeptin Neurons
Estradiol exerts opposite effects on the two kisspeptin neuron populations. Positive feedback on AVPV neurons and negative feedback on arcuate neurons. In the days leading to ovulation, rising estradiol levels activate ERα in AVPV kisspeptin neurons, increasing KISS1 mRNA expression and kisspeptin peptide content. At a threshold estradiol concentration (~200 pg/mL sustained for 36–48 hours in humans), AVPV kisspeptin neurons trigger a massive, sustained burst of GnRH secretion lasting 12–24 hours. The GnRH surge that drives the preovulatory LH surge. Blocking kisspeptin signaling with GPR54 antagonists during this window abolishes the LH surge and prevents ovulation, even when estradiol levels are optimal.
In contrast, arcuate kisspeptin neurons express both ERα and ERβ, with ERα mediating negative feedback. Low estradiol (early follicular phase or after ovariectomy) increases KISS1 expression in ARC neurons, driving higher GnRH pulse frequency. The signal that stimulates FSH secretion and follicle recruitment. High estradiol suppresses ARC kisspeptin, slowing GnRH pulse frequency and reducing LH secretion during the luteal phase. This dual regulatory system allows a single steroid (estradiol) to coordinate both surge generation and pulse frequency modulation through spatially distinct kisspeptin neuron populations.
Testosterone and dihydrotestosterone (DHT) act similarly in males, with negative feedback on arcuate kisspeptin neurons maintaining stable LH pulsatility. Castration causes KISS1 mRNA in the arcuate nucleus to increase 5–10-fold within one week, and LH levels rise proportionally. Testosterone replacement suppresses arcuate kisspeptin back to baseline, normalizing LH. Interestingly, aromatization of testosterone to estradiol is required for full feedback. Non-aromatizable androgens like DHT produce weaker suppression of kisspeptin expression, indicating that ERα mediates much of the negative feedback even in males.
Kisspeptin neuron populations are the mechanistic interface where gonadal steroids communicate reproductive status back to the brain. They translate circulating hormone concentrations into changes in GnRH neuron firing patterns. Without this feedback loop, the HPG axis would operate in an open-loop mode, unable to adjust output based on gonadal response.
The complexity here matters. Researchers designing fertility models or studying pubertal timing cannot treat 'estrogen' or 'testosterone' as monolithic variables. The effect depends entirely on which kisspeptin neuron population you're looking at, which receptor subtype is expressed, and what the temporal pattern of steroid exposure has been. A single bolus of estradiol produces different outcomes than sustained elevation over 48 hours because the transcriptional machinery in AVPV neurons requires time to accumulate kisspeptin peptide before surge capacity is reached.
If your research question touches reproductive neuroendocrinology or metabolic-reproductive interactions, precise peptide tools are non-negotiable. Contaminant peptides with off-target receptor activity or degraded sequences will produce results that look like 'low responders' or 'heterogeneous populations' when the real issue is peptide quality. Explore high-purity research peptides synthesized with exact amino-acid sequencing to ensure every experiment reflects endogenous signaling fidelity. Not synthesis artifacts.
Kisspeptin signaling is the obligate gatekeeper of mammalian reproduction. Disrupt it and puberty never starts. Block it in adulthood and fertility ceases within days. Understanding this pathway at the molecular level isn't academic curiosity. It's the foundation for every experimental model studying reproductive timing, metabolic influences on fertility, or neuroendocrine regulation of the HPG axis. The system is elegant, essential, and unforgiving of imprecision.
Frequently Asked Questions
What is the kisspeptin signaling pathway and why is it essential for reproduction?▼
The kisspeptin signaling pathway is a neuroendocrine cascade where kisspeptin neurons in the hypothalamus release kisspeptin peptides that bind to GPR54 receptors on GnRH neurons, triggering GnRH secretion. GnRH then stimulates the anterior pituitary to release LH and FSH, which regulate gonadal function. This pathway is essential because loss-of-function mutations in the KISS1 gene or GPR54 receptor cause hypogonadotropic hypogonadism — a condition where puberty fails to initiate and fertility never develops. Kisspeptin is the obligate upstream activator of the entire reproductive hormone axis — without it, GnRH neurons remain silent despite being anatomically normal.
How does kisspeptin activate GnRH neurons at the molecular level?▼
Kisspeptin binds to GPR54, a Gq-coupled GPCR on the surface of GnRH neurons, activating phospholipase C which generates IP3 and DAG. IP3 mobilizes calcium from intracellular stores, depolarizing the neuron membrane and triggering action potentials that drive GnRH secretion. Electrophysiological studies show that kisspeptin increases GnRH neuron firing frequency from a basal 0.1 Hz to 2–3 Hz within 60 seconds of application. This calcium-dependent mechanism is direct — kisspeptin does not require intermediary neurotransmitters or secondary cell types to activate GnRH neurons.
What happens if kisspeptin signaling is blocked in adult animals?▼
Blocking kisspeptin signaling with GPR54 antagonists in adult animals causes immediate suppression of LH pulsatility within 2–4 hours, with pulse frequency dropping to near zero. Continuous antagonism over seven days produces gonadal atrophy comparable to surgical castration — testes weight decreases by 40–50%, spermatogenesis arrests, and estrous cycles cease in females. Serum LH and sex steroid levels fall to prepubertal ranges within one week. The effect is reversible — LH pulsatility resumes within 12–24 hours after antagonist washout, and fertility is restored within 4–6 weeks as gametogenesis reinitializes.
Why do researchers studying reproductive biology need high-purity kisspeptin peptides?▼
Kisspeptin-10 has a plasma half-life under 30 minutes due to peptidase degradation, and even minor amino-acid substitutions at the C-terminal arginine residues abolish GPR54 binding. Contaminant peptides or degraded sequences produce inconsistent GnRH responses that appear as ‘low responders’ or ‘heterogeneous populations’ when the real issue is peptide quality. Research-grade kisspeptin requires exact amino-acid sequencing verified by mass spectrometry to replicate endogenous signaling fidelity. Protocols using peptides with >95% purity and immediate post-reconstitution administration produce reproducible activation of GnRH neurons and LH secretion across experiments.
How do kisspeptin neurons integrate metabolic signals like leptin?▼
Kisspeptin neurons in the arcuate nucleus express leptin receptors, and leptin binding activates JAK-STAT signaling pathways that upregulate KISS1 gene transcription. When energy availability is high and leptin levels are sufficient, kisspeptin expression increases and the reproductive axis remains active. During negative energy balance (caloric restriction, low body fat), leptin drops, kisspeptin expression falls, and GnRH secretion is suppressed — effectively shutting down reproduction to preserve survival. Leptin-deficient ob/ob mice are infertile with undetectable LH, but leptin administration restores KISS1 expression within 24 hours and fertility within two weeks, demonstrating that kisspeptin neurons serve as the metabolic gate for reproduction.
What is the difference between AVPV and arcuate kisspeptin neuron populations?▼
AVPV kisspeptin neurons mediate the preovulatory GnRH and LH surge in females by responding to rising estradiol with increased kisspeptin expression and sustained GnRH secretion lasting 12–24 hours. Arcuate nucleus kisspeptin neurons (KNDy neurons) generate pulsatile GnRH secretion in both sexes through synchronized bursting driven by co-released neurokinin B (excitatory) and dynorphin (inhibitory), producing ~60-minute pulse intervals. AVPV neurons are primarily involved in ovulation triggering, while arcuate neurons regulate baseline reproductive axis activity and pulse frequency modulation throughout the cycle.
Can exogenous kisspeptin administration restore fertility in hypogonadal patients?▼
Intravenous kisspeptin-54 administration (0.24–1.0 nmol/kg) in men with idiopathic hypogonadotropic hypogonadism stimulates LH secretion within 30 minutes and raises testosterone by 20–30% within 2–4 hours. However, the effect is transient — LH returns to baseline within 6–8 hours, and repeated daily dosing causes tachyphylaxis within one week due to GPR54 receptor desensitization. This contrasts with pulsatile GnRH therapy, which maintains efficacy over months. Current research is exploring kisspeptin analogs with modified structures that resist degradation and maintain pulsatile signaling without receptor downregulation, but no formulations have reached clinical use.
How does estradiol feedback regulate kisspeptin neurons differently in AVPV vs arcuate populations?▼
Estradiol exerts positive feedback on AVPV kisspeptin neurons via ERα, increasing KISS1 expression when estradiol is sustained above ~200 pg/mL for 36–48 hours — this triggers the preovulatory GnRH surge. In contrast, estradiol exerts negative feedback on arcuate kisspeptin neurons, suppressing KISS1 expression during high-estradiol phases like the luteal phase and slowing GnRH pulse frequency. Low estradiol (early follicular phase) increases arcuate kisspeptin expression and GnRH pulse frequency, driving FSH secretion for follicle recruitment. This dual regulation allows the same steroid to coordinate both surge generation and pulse frequency modulation through spatially distinct neuron populations.
What role does neurokinin B play in the kisspeptin signaling pathway?▼
Neurokinin B (NKB) is co-released with kisspeptin from arcuate KNDy neurons and acts as an autoexcitatory signal by binding NK3 receptors on neighboring KNDy neurons, propagating synchronized bursts across the network. This creates recurrent excitation-inhibition cycles when dynorphin terminates each burst, generating the ~60-minute GnRH pulse intervals required for normal reproductive function. Mutations in TAC3 (encoding NKB) or TACR3 (encoding NK3 receptor) cause hypogonadotropic hypogonadism, and NK3 receptor antagonists block LH pulsatility in adult animals, demonstrating that NKB is an essential component of the GnRH pulse generator — not a modulatory signal.
Why do protocols using kisspeptin peptides sometimes produce inconsistent results?▼
Kisspeptin-10 has a plasma half-life under 30 minutes due to peptidase degradation at the N-terminus, and peptide solutions that sit at room temperature for more than 30–60 minutes lose significant bioactivity. Protocols that reconstitute peptides and then delay administration, use peptides with <95% purity, or fail to account for dose-response threshold behavior (minimal effect below 0.3 nmol/kg in rodents) produce inconsistent GnRH neuron activation and LH responses. Additionally, kisspeptin neurons operate as a network with NKB and dynorphin co-transmission — isolating kisspeptin effects without accounting for this integrated signaling often yields results that appear variable when the real issue is incomplete understanding of the endogenous system.











