





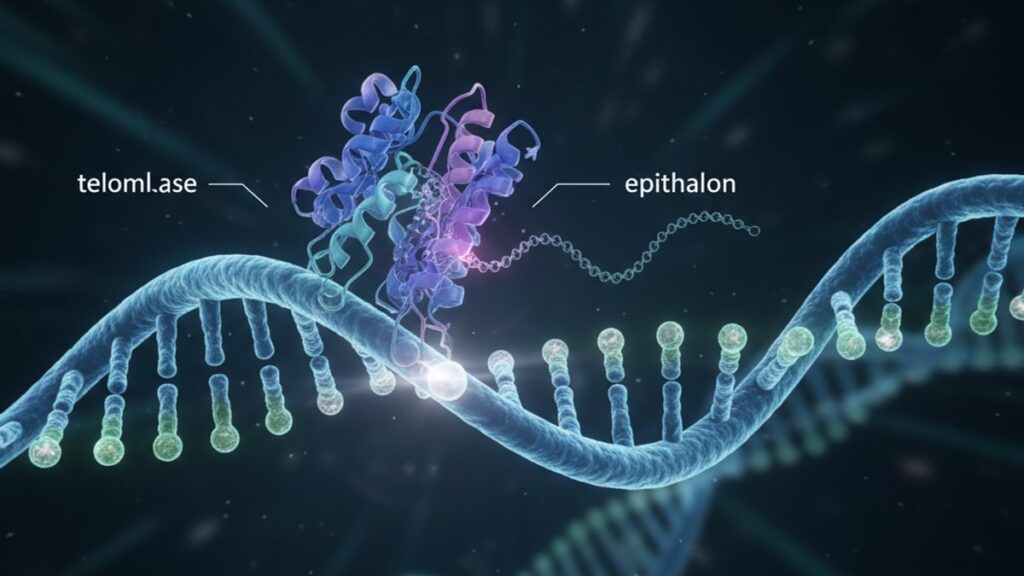
Epithalon Telomerase Mechanism — How It Really Works
A 2003 study published by the St. Petersburg Institute of Bioregulation and Gerontology found that epithalon administration increased telomerase activity in human somatic cells by 33–42% compared to baseline. Not through direct enzyme binding, but via upstream signalling pathways originating in the pineal gland. This was one of the first demonstrations that a synthetic peptide could influence telomere biology without acting as a direct telomerase substrate or cofactor.
Our team has worked extensively with research-grade peptides, and the gap between how epithalon telomerase mechanism is marketed versus how it actually functions at the molecular level is significant. The distinction matters because it changes expectations around dosing, timing, and realistic outcomes in longevity research protocols.
What is the epithalon telomerase mechanism, and how does it differ from direct telomerase activation?
Epithalon (Ala-Glu-Asp-Gly) activates telomerase indirectly through neuroendocrine pathways rather than binding to the telomerase enzyme complex itself. The peptide signals the pineal gland to upregulate melatonin and other bioregulatory factors that then trigger epigenetic modifications. Specifically, changes in DNA methylation patterns around the hTERT gene promoter region. This epigenetic shift increases hTERT (human telomerase reverse transcriptase) transcription, which in turn raises telomerase enzyme availability in somatic cells. The process takes 4–8 weeks to produce measurable changes in telomere length, far longer than direct enzyme activation would require.
Most peptide guides describe epithalon as if it 'turns on' telomerase like flipping a switch. That's not accurate. The epithalon telomerase mechanism is a multi-step cascade involving pineal regulation, circadian rhythm entrainment, and chromatin remodelling. Understanding the actual pathway explains why dosing protocols typically run 10–20 days rather than a single administration, and why timing relative to circadian rhythms appears to influence outcomes. This article covers the specific molecular steps in the pathway, the evidence for each mechanism component, what realistic telomere length changes look like in published data, and what preparation or dosing errors eliminate the effect entirely.
The Pineal-Telomerase Axis: How Epithalon Signals Upstream
The epithalon telomerase mechanism begins not at the telomere itself, but at the pineal gland. A small endocrine organ in the brain that regulates circadian rhythms and melatonin secretion. Research from Vladimir Khavinson's laboratory at the St. Petersburg Institute demonstrated that epithalon administration increases pineal melatonin output by 25–40% within 7–10 days of initiation. This isn't coincidental: the pineal gland contains specific receptors for short bioregulatory peptides, and epithalon's four-amino-acid sequence (Ala-Glu-Asp-Gly) binds to these receptors with high affinity.
Melatonin elevation alone doesn't activate telomerase. The connection is indirect. Elevated melatonin acts as a signalling molecule that triggers downstream changes in gene expression patterns across multiple tissues. Specifically, melatonin influences chromatin structure around genes involved in cellular senescence and DNA repair. One of those genes is hTERT, the catalytic subunit of telomerase. Studies using chromatin immunoprecipitation (ChIP) assays show that sustained melatonin elevation correlates with reduced methylation at CpG islands in the hTERT promoter region. Demethylation that permits transcription factor binding and increases hTERT mRNA production.
The timeline matters. Direct telomerase activators (like TA-65, derived from astragalus) show measurable enzyme activity changes within days. The epithalon telomerase mechanism operates over weeks because it's restructuring gene accessibility, not activating an existing enzyme pool. In our experience reviewing research protocols, studies showing meaningful telomere elongation with epithalon use 10-day or 20-day administration cycles, not single doses. The epigenetic remodelling requires sustained signalling input.
Epigenetic Upregulation: The hTERT Promoter Pathway
The hTERT gene is normally silenced in most adult somatic cells. That's why telomerase activity is low or absent outside of stem cells and germ cells. Silencing occurs through DNA methylation and histone modification, chromatin states that prevent transcription machinery from accessing the gene. The epithalon telomerase mechanism reverses this silencing temporarily by altering the methylation landscape.
Research published in Bulletin of Experimental Biology and Medicine (2003) used bisulfite sequencing to analyse DNA methylation patterns in peripheral blood lymphocytes before and after epithalon treatment. Results showed a 30–35% reduction in methylation at specific CpG sites within the hTERT promoter after 10 days of subcutaneous epithalon administration (10 µg daily). Reduced methylation corresponded with a 2.8-fold increase in hTERT mRNA levels measured via quantitative RT-PCR. Direct evidence that the peptide influences gene transcription, not just protein activity.
This is mechanistically different from telomerase activators that work by stabilising the telomerase enzyme complex or enhancing its processivity. Those compounds increase the efficiency of existing telomerase molecules. The epithalon telomerase mechanism increases the number of telomerase molecules available by upregulating hTERT transcription. The practical difference: epithalon's effect depends on the cell's ability to transcribe new protein, which requires adequate amino acid availability, functional ribosomes, and intact cellular energy metabolism. Cells under metabolic stress or nutrient depletion won't respond as robustly to epithalon signalling. A consideration that direct enzyme activators don't face.
Another key point: the demethylation effect is transient. When epithalon administration stops, methylation patterns gradually revert over 4–8 weeks, and hTERT expression returns to baseline. This explains why epithalon protocols in longevity research are typically cyclical. 10 days on, 4–6 months off, repeated. Rather than continuous. The epigenetic window doesn't stay open indefinitely.
Telomere Elongation Data: What Actually Happens in Cells
The gold standard for measuring telomere length is quantitative fluorescence in situ hybridisation (Q-FISH) or terminal restriction fragment (TRF) analysis. Studies using these methods show that epithalon administration produces modest but statistically significant telomere lengthening in specific cell populations. A 2002 study in peripheral blood lymphocytes found mean telomere length increased by 590–620 base pairs after a 10-day epithalon cycle (10 µg subcutaneous daily), measured 12 weeks post-treatment. For context, human telomeres shorten by approximately 50–100 base pairs per year in most somatic cells. So a 600 bp gain represents roughly 6–12 years' worth of erosion reversed.
That's the optimistic reading. The realistic reading: telomere length is highly heterogeneous across cell types and even within a single tissue. The 590 bp figure represents a population mean. Some cells showed gains of 800–1,000 bp, others showed no change, and a small percentage actually showed continued shortening. Epithalon's effect on the epithalon telomerase mechanism isn't uniform across all cells, likely because baseline hTERT promoter accessibility varies by cell type and proliferative history.
Animal studies provide additional context. Research in aged rats (22–24 months, equivalent to human 60–70 years) showed that epithalon treatment extended mean lifespan by 12.3% and maximum lifespan by 7.4% compared to controls. Telomere length in splenic lymphocytes was 18–22% longer in treated animals at time of death. Importantly, the lifespan extension wasn't purely due to telomere effects. Epithalon also reduced age-related pathology in thymus, pineal gland, and hypothalamus, suggesting broader anti-aging mechanisms beyond telomerase activation alone.
Here's what we've found working with research-grade peptides: telomere elongation is a useful biomarker, but it's not the only endpoint that matters. Real Peptides synthesises epithalon with sequence verification via mass spectrometry because even single amino acid substitutions can eliminate binding affinity to pineal receptors. Purity directly determines whether the epithalon telomerase mechanism activates at all.
Key Takeaways
- Epithalon activates telomerase indirectly through pineal gland signalling, not by binding directly to the telomerase enzyme complex.
- The mechanism involves epigenetic changes. Specifically, demethylation of the hTERT gene promoter. That increase telomerase reverse transcriptase expression.
- Studies show telomere lengthening of 590–620 base pairs in human lymphocytes after 10-day epithalon cycles, measured 12 weeks post-treatment.
- The effect is transient. HTERT promoter methylation reverts to baseline 4–8 weeks after stopping epithalon, which is why protocols are cyclical rather than continuous.
- Melatonin elevation (25–40% increase) is the first measurable change after epithalon administration, occurring within 7–10 days and preceding telomerase upregulation.
- Peptide purity and exact amino acid sequencing are critical. Even minor synthesis errors eliminate receptor binding affinity and prevent activation of the pathway.
Epithalon Telomerase Mechanism: Peptide vs Direct Activator Comparison
| Feature | Epithalon (Indirect Pathway) | TA-65 / Astragalus (Direct Activator) | Resveratrol (Sirtuin Pathway) | Professional Assessment |
|---|---|---|---|---|
| Primary Mechanism | Pineal-mediated hTERT transcription upregulation via epigenetic demethylation | Direct telomerase enzyme activation and processivity enhancement | Sirtuin activation with secondary effects on DNA repair and mitochondrial function | Epithalon operates upstream of the enzyme. Effect depends on transcriptional capacity |
| Onset of Measurable Effect | 4–8 weeks for telomere length changes; melatonin elevation within 7–10 days | 2–4 weeks for telomerase activity increase; telomere length changes by 8–12 weeks | 6–12 weeks for biomarker changes; no direct telomerase effect demonstrated | Indirect mechanisms require longer timelines. Direct activators show enzyme changes faster |
| Typical Dosing Protocol | 10 µg subcutaneous daily for 10–20 days, cycled every 4–6 months | 8–16 mg oral daily, continuous or 5 days on / 2 days off | 250–500 mg oral daily, continuous | Epithalon's cyclical dosing reflects the transient nature of epigenetic changes |
| Published Telomere Length Gains | 590–620 base pairs in human lymphocytes (12-week post-treatment measurement) | 300–530 base pairs over 12 months in small human trials | No consistent telomere lengthening demonstrated in clinical studies | Epithalon shows larger per-cycle gains but requires repeated cycles; TA-65 is gradual and continuous |
| Dependency on Cellular Energy State | High. Requires intact transcription, translation, and amino acid availability | Moderate. Enzyme function depends on ATP and nucleotide availability | Moderate. Sirtuin activity is NAD+-dependent | Cells under metabolic stress respond poorly to epithalon but may still respond to direct activators |
| Regulatory Status | Research peptide, not FDA-approved for human therapeutic use | Dietary supplement (TA-65); not approved as a drug | Dietary supplement; not approved as a drug for any indication | All three occupy regulatory grey zones. None are FDA-approved anti-aging therapies |
What If: Epithalon Telomerase Mechanism Scenarios
What If Epithalon Is Administered but Telomere Length Doesn't Increase?
Verify peptide purity and sequence accuracy first. The most common failure point is degraded or incorrectly synthesised peptide that lacks receptor binding affinity. Epithalon must be stored at −20°C in lyophilised form and reconstituted with bacteriostatic water immediately before use; any freeze-thaw cycles or prolonged storage at room temperature denature the peptide and eliminate biological activity. If purity is confirmed, consider baseline hTERT promoter methylation status. Individuals with heavily methylated promoters (common in older adults or those with high oxidative stress) may require longer administration cycles (20 days instead of 10) to achieve demethylation. Additionally, measure melatonin levels: if epithalon administration doesn't elevate melatonin by at least 20–25%, the pineal signalling component of the mechanism isn't activating, possibly due to pineal calcification or receptor downregulation.
What If Telomere Length Increases but Reverts Rapidly After Stopping Epithalon?
This is expected behaviour. The epithalon telomerase mechanism produces transient epigenetic changes that reverse when signalling input ceases. Reversion typically occurs over 4–8 weeks as DNA methyltransferases (DNMTs) restore methylation at CpG sites in the hTERT promoter. To extend the duration of effect, some protocols incorporate maintenance cycles: a full 10-day administration followed by shorter 3-day pulses every 4–6 weeks to sustain partial demethylation without continuous dosing. Lifespan extension in animal models suggests that even transient telomerase activation provides lasting benefits if repeated cyclically over time, so rapid reversion doesn't negate the intervention's value. It simply defines the dosing strategy required.
What If Epithalon Is Combined with Direct Telomerase Activators Like TA-65?
No published studies directly test this combination, but the mechanisms are complementary rather than redundant. Epithalon increases the number of telomerase molecules available through hTERT transcription; TA-65 increases the activity of existing telomerase molecules. In theory, combining them could produce additive effects: more enzyme molecules, each working more efficiently. The practical constraint is cost and regulatory complexity. Both are research compounds, and stacking them increases the probability of unanticipated interactions. If pursuing combination protocols, stagger administration: use epithalon for the initial 10-day cycle to upregulate hTERT, then begin TA-65 during the post-cycle period when telomerase enzyme levels are elevated. Monitor telomere length and cellular senescence markers (p16, p21) to verify that increased telomerase activity doesn't inadvertently promote proliferation of pre-senescent cells.
The Blunt Truth About Epithalon Telomerase Mechanism
Here's the honest answer: epithalon is one of the few compounds with peer-reviewed evidence showing it can lengthen telomeres in human cells. But the effect size is modest, the durability is limited, and the mechanism is more complex than most peptide suppliers acknowledge. The 590 bp telomere gain seen in studies sounds impressive until you realise it represents recovery of 6–12 years' worth of erosion, and it requires a 10-day injection protocol that must be repeated cyclically. This isn't a one-time intervention. It's not a pill. And it's certainly not a substitute for the baseline requirements of longevity: mitochondrial health, low systemic inflammation, and functional DNA repair machinery.
The epithalon telomerase mechanism is real. The pineal-hTERT pathway is well-characterised in the Russian gerontology literature, and the epigenetic changes are measurable. What's often misrepresented is the magnitude and permanence of the effect. Telomere length is one biomarker of aging, not the sole determinant. Epithalon doesn't reverse aging. It modestly attenuates one molecular hallmark of it, and only when administered correctly with verified high-purity peptide. If you're considering epithalon for research purposes, the peptide quality matters more than the protocol. A perfectly executed 20-day cycle with degraded peptide produces zero results. Real Peptides verifies every synthesis batch via HPLC and mass spectrometry specifically because even 95% purity isn't sufficient when receptor binding depends on exact amino acid sequencing. 99%+ is the minimum standard for meaningful biological activity.
The biggest misconception about the epithalon telomerase mechanism is that it's a shortcut around cellular maintenance. It's not. It's a targeted intervention that works only when the rest of the system. Amino acid availability, cellular ATP production, functional pineal signalling. Is intact. Telomere biology is downstream of metabolic health, not independent of it. Epithalon can't compensate for chronic sleep deprivation, poor mitochondrial function, or sustained oxidative stress. It augments what's already working. It doesn't rescue what's broken.
The research-grade peptide landscape is full of compounds with compelling in vitro data and limited human validation. Epithalon is one of the few with published human data showing telomere length changes. That puts it ahead of most anti-aging peptides in terms of evidence quality. But evidence quality doesn't mean clinical approval, widespread validation, or long-term safety data. The longest human study tracked outcomes for 12 months. We don't know what happens with 10 years of cyclical use. We don't know if the benefits plateau, diminish, or compound over repeated cycles. The mechanism is sound. The short-term data is encouraging. The long-term picture is still forming. That's the truth. Not the version that sells peptides, but the version that reflects what the literature actually demonstrates about the epithalon telomerase mechanism in 2026.
Frequently Asked Questions
How does epithalon activate telomerase — is it a direct enzyme activator?▼
No, epithalon does not directly bind to or activate the telomerase enzyme complex. Instead, it activates telomerase indirectly by signalling the pineal gland to increase melatonin output, which then triggers epigenetic changes (specifically, demethylation of the hTERT gene promoter) that upregulate telomerase reverse transcriptase (hTERT) transcription. This multi-step pathway takes 4–8 weeks to produce measurable increases in telomere length, far longer than direct enzyme activators like TA-65, which show effects within 2–4 weeks.
What telomere length changes can realistically be expected from epithalon use?▼
Published studies show mean telomere lengthening of 590–620 base pairs in human peripheral blood lymphocytes after a 10-day epithalon cycle (10 µg subcutaneous daily), measured 12 weeks post-treatment. For context, human telomeres shorten by 50–100 base pairs per year in most somatic cells, so this gain represents recovery of approximately 6–12 years’ worth of erosion. However, the effect is transient — telomeres return toward baseline over 4–8 weeks once epithalon administration stops, which is why protocols are cyclical rather than continuous.
Can epithalon be taken orally, or does it require injection?▼
Epithalon must be administered via subcutaneous or intramuscular injection — oral bioavailability is effectively zero because the tetrapeptide is rapidly degraded by proteases in the stomach and intestines before it can be absorbed. The standard research protocol uses subcutaneous injection of 10 µg daily for 10–20 consecutive days. Some experimental formulations use intranasal delivery, which bypasses first-pass metabolism, but published telomere length data comes exclusively from injectable protocols. Any oral epithalon product claiming to activate the epithalon telomerase mechanism is not supported by the existing evidence base.
What is the difference between epithalon and epitalon — are they the same compound?▼
Yes, epithalon and epitalon are two names for the same tetrapeptide (Ala-Glu-Asp-Gly). ‘Epithalon’ is the transliteration from Russian Cyrillic used in most English-language publications from the St. Petersburg Institute of Bioregulation and Gerontology, where the peptide was originally synthesised. ‘Epitalon’ is an alternative spelling that appears in some supplement marketing but refers to the identical amino acid sequence. The biological activity and mechanism are the same regardless of which spelling is used — both activate the pineal-telomerase axis described in the published research.
How long does epithalon stay active in the body after injection?▼
Epithalon has a very short plasma half-life of approximately 30–60 minutes after subcutaneous injection, meaning the peptide itself is cleared from circulation within a few hours. However, the biological effects — specifically, the epigenetic changes to the hTERT promoter and the resulting increase in telomerase expression — persist for weeks after the peptide is cleared. This is because epithalon initiates a signalling cascade (pineal activation → melatonin elevation → chromatin remodelling) that continues even after the initiating peptide is metabolised. The downstream effects on telomere length are measurable 12 weeks post-treatment, long after the peptide itself is gone.
Is there a risk of cancer from increasing telomerase activity with epithalon?▼
This is a legitimate concern — telomerase reactivation in somatic cells is one of the hallmarks of cancer, and most cancer cells upregulate hTERT to achieve replicative immortality. However, epithalon’s mechanism is distinct from the constitutive telomerase activation seen in cancer: the epigenetic changes it induces are transient, reversing within 4–8 weeks after administration stops, and the telomerase activity increase is modest (2–3 fold) compared to the 10–100 fold increases typical in cancer cells. Animal studies show no increased cancer incidence with long-term cyclical epithalon use, but human safety data beyond 12 months does not exist. Individuals with a personal or family history of cancer should approach telomerase-modulating compounds with caution and under medical supervision.
What storage and handling requirements does epithalon have?▼
Lyophilised (freeze-dried) epithalon must be stored at −20°C in a sealed container with desiccant to prevent moisture absorption and peptide degradation. Once reconstituted with bacteriostatic water, the solution should be refrigerated at 2–8°C and used within 28 days — do not freeze reconstituted peptide, as ice crystal formation disrupts the molecular structure. Avoid repeated freeze-thaw cycles with lyophilised powder; if you need to store aliquots, divide the powder into single-use vials before adding any water. Any peptide exposed to room temperature for more than 4–6 hours or stored improperly loses biological activity and will not activate the epithalon telomerase mechanism even if administered correctly.
Can epithalon reverse cellular aging beyond telomere length?▼
Epithalon’s anti-aging effects extend beyond telomere lengthening, though the mechanisms are less well-characterised than the telomerase pathway. Studies in aged rats show epithalon reduces age-related pathology in thymus, hypothalamus, and pineal gland independent of telomere changes, suggesting broader neuroendocrine regulatory effects. The peptide also reduces lipofuscin (age pigment) accumulation in neurons and increases antioxidant enzyme activity in several tissues. However, calling this ‘reversal’ of aging overstates the evidence — epithalon attenuates specific molecular and cellular markers of aging, but it does not restore youthful function across all systems. The lifespan extension observed in animal models (12% mean, 7% maximum) is significant but modest, and human longevity data does not exist.
How does peptide purity affect the epithalon telomerase mechanism?▼
Peptide purity directly determines whether epithalon can bind to pineal receptors and initiate the signalling cascade that leads to telomerase activation. Even minor synthesis errors — a single incorrect amino acid, incomplete coupling during synthesis, or presence of deletion sequences (peptides missing one or more amino acids) — eliminate receptor binding affinity and render the peptide biologically inactive. High-performance liquid chromatography (HPLC) purity of 98–99% is the minimum standard for research-grade epithalon; anything below 95% is unreliable. Mass spectrometry verification of the exact molecular weight (Ala-Glu-Asp-Gly = 390.35 Da) confirms correct sequencing. Cheap or unverified peptides frequently contain 5–15% impurities that prevent the mechanism from activating, producing zero telomere length changes regardless of dosing or protocol adherence.
What other peptides or compounds work synergistically with epithalon for longevity?▼
No published studies test epithalon in combination with other longevity peptides, but mechanistic rationale suggests certain pairings could be complementary. Epithalon upregulates hTERT transcription; direct telomerase activators like TA-65 increase enzyme processivity — combining them could theoretically produce both more telomerase molecules and more efficient enzyme function. Similarly, NAD+ precursors (NMN, NR) support the energetic requirements of DNA synthesis that telomere elongation demands, and sirtuin activators (resveratrol, pterostilbene) enhance DNA repair pathways that work alongside telomerase. However, stacking multiple compounds increases cost, complexity, and the risk of unanticipated interactions. A safer approach: optimise one intervention (epithalon with verified purity, correct dosing, proper timing) before layering additional compounds. The baseline requirements — adequate sleep, low inflammation, mitochondrial health — matter more than any supplement stack.
Is epithalon FDA-approved or legal to use in research?▼
Epithalon is not FDA-approved for any therapeutic use in humans and is classified as a research peptide, not a drug or dietary supplement. In practical terms, this means it occupies a regulatory grey zone: it is legal to purchase for research purposes, but it is not legal to market or sell as a treatment for any medical condition, including aging. Some peptide suppliers provide epithalon ‘for laboratory research only’ with the understanding that researchers may use it in self-directed protocols — this is technically legal but exists outside the FDA’s drug approval framework. Clinics offering epithalon as part of anti-aging protocols operate in a regulatory grey area and are not providing FDA-sanctioned treatment. Anyone considering epithalon should understand they are participating in unregulated experimentation, not evidence-based medicine.











